GRAB Sensors - Tools for Precisely Measuring Neurochemical Dynamics in vivo
1. Development Principle of GRAB Sensors
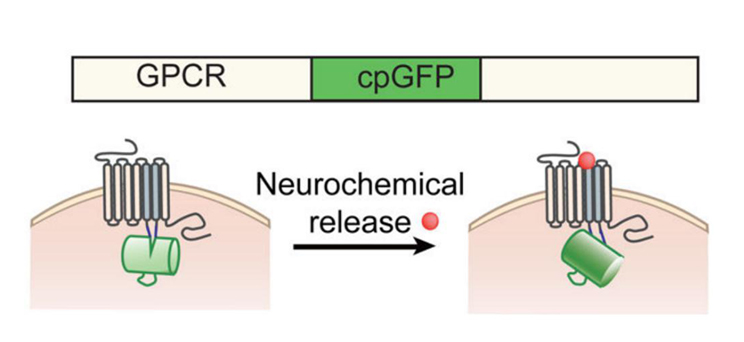
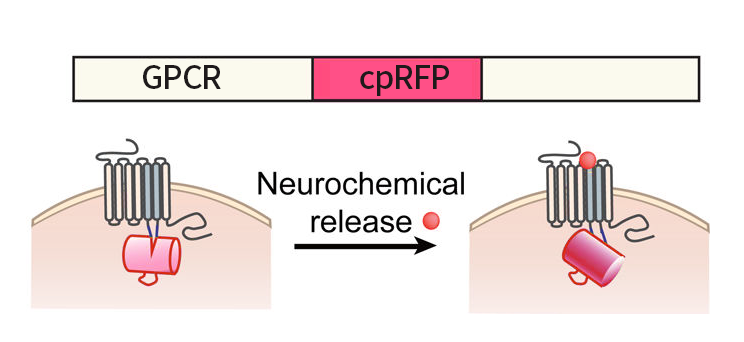
Through the efforts the Yulong Li lab in Peking University, many genetically-modified fluorescent sensors delivered by AAVs for visualizing specific neurochemicals were developed by coupling neurochemical-sensing G protein coupled receptors (GPCRs) with a circularly permutated fluorescent protein (cpFP). When neurochemicals bind to their specific GPCRs, the conformation change in these GPCRs gives rise to a change in fluorescence of cpFPs, which can be detected by fluorescence imaging (Fig 1). These sensors are named as GRAB (GPCR Activation-Based) sensors, which can be used to in vivo monitor the dynamic changes of these neurochemicals with high spatiotemporal resolutions.
Fig 1: Schematic drawing shows the principle of the green and red GRAB sensors. Upon binding its ligand, the conformational change in the GPCR is sensed and reported by cpGFP or cpRFP (modified based on reference 4).
2. Properties of Selected Published Sensor

|
Sensor
|
Neurochemical Measured
|
Version
|
Color
|
Scaffold
|
Affinity
|
Peak Response
(∆F/F0)
|
Kinetics
|
Coupling to Downstream Signaling Pathway
|
Reference
|

|
Ach2.0
|
Acetylcholine
|
First generation
|
Green
|
Human M3 receptor
|
EC50~1uM
|
~90%
|
τon ~200ms,
τoff~ 800ms
|
Weak coupling
|
[8]
|

|
Ach3.0
|
Acetylcholine
|
Second generation
|
Green
|
Human M3 receptor
|
EC50~2uM
|
~280%
|
τon ~112ms,
τoff~ 580ms
|
Negligible coupling
|
[3]
|

|
Ach3.0-mut
|
Acetylcholine
|
Second generation
control
|
Green
|
Human M3 receptor
|
EC50~0uM
(W200A mutation)
|
~1.8%
|
-
|
-
|
[3]
|
|
|
DA1m
|
Dopamine
|
First generation
|
Green
|
Human D2 receptor
|
EC50~130nM
Medium affinity
|
~90%
|
τon~60ms,
τoff~700ms
|
Negligible coupling
|
[7]
|
|
|
DA1h
|
Dopamine
|
First generation
|
Green
|
Human D2 receptor
|
EC50~10nM
High affinity
|
~90%
|
τon~140ms,
τoff~2500ms
|
Negligible coupling
|
[7]
|
|
|
DAmut
(1st)
|
Dopamine
|
First generation
control
|
Green
|
Human D2 receptor
|
EC50~0uM
C118A & S193N mutations
|
No effect
|
-
|
-
|
[7]
|
|
|
DA2m(DA4.4)
|
Dopamine
|
Second generation
|
Green
|
Human D2 receptor
|
EC50~90nM
Medium affinity
|
~340%
|
τon ~40ms,
τoff~1300ms
|
Minimal coupling
|
[2]
|
|
|
DA2h(DA4.3)
|
Dopamine
|
Second generation
|
Green
|
Human D2 receptor
|
EC50~7nM
High affinity
|
~280%
|
τon ~50ms,
τoff~ 7300ms
|
Minimal coupling
|
[2]
|
|
|
DAmut(2nd)
|
Dopamine
|
Second generation
control
|
Green
|
Human D2 receptor
|
EC50~0uM
C1183.36A & S1935.42N mutations
|
No effect
|
-
|
-
|
[2]
|
|
|
rDA1m (rDA2.5m)
|
Dopamine
|
-
|
Red
|
Human D2 receptor
|
EC50~95nM
medium-affinity
|
~150%
|
τon ~80ms,
τoff~ 770ms
|
Minimal coupling
|
[2]
|
|
|
rDA1h (rDA2.5h)
|
Dopamine
|
-
|
Red
|
Human D2 receptor
|
EC50~4nM
high-affinity
|
~100%
|
τon ~60ms,
τoff~ 2150ms
|
Minimal coupling
|
[2]
|
|
|
rDAmut (rDA2.5mut)
|
Dopamine
|
Control
|
Red
|
Human D2 receptor
|
EC50~0uM
C1183.36A & S1935.42N mutations
|
No effect
|
-
|
-
|
[2]
|
|
|
NE1m(NE2.1)
|
NE
|
-
|
Green
|
Human a2A receptor
|
EC50~930nM
Medium affinity
|
~230%
|
τon ~72ms,
τoff~680ms
|
No coupling
|
[6]
|
|
|
NE1h(NE2.2)
|
NE
|
-
|
Green
|
Human a2A receptor
|
EC50~83nM
High affinity
|
~130%
|
τon ~36ms,
τoff~1890ms
|
No coupling
|
[6]
|
|
|
NEmut
|
NE
|
Control
|
Green
|
Human a2A receptor
|
EC50~0uM
S5.46A mutation
|
No effect
|
-
|
-
|
[6]
|
|
|
Ado1.0
|
Adenosine
|
-
|
Green
|
Human A2A receptor
|
EC50~60nM
|
~130%
|
τon~36ms,
τoff~1890ms
|
Negligible coupling
|
[4]
|
|
|
Ado1.0mut
|
Adenosine
|
Control
|
Green
|
Human A2A receptor
|
EC50~0uM
F168A mutation
|
No effect
|
-
|
-
|
[4]
|
|
|
5-HT1.0
|
Serotonin
|
-
|
Green
|
Human 5-HT2C receptor
|
EC50~22nM
|
~250%
|
τon~200ms,
τoff~3100ms
|
No coupling
|
[1]
|
|
|
5-HTmut
|
Serotonin
|
Control
|
Green
|
Human 5-HT2C receptor
|
EC50~0uM
D1343.32Q mutation
|
No effect
|
-
|
-
|
[1]
|
Please note that most of the properties mentioned in the above are from measurements in vitro in cultured cells expressing the GRAB sensors.
3. How to Order?
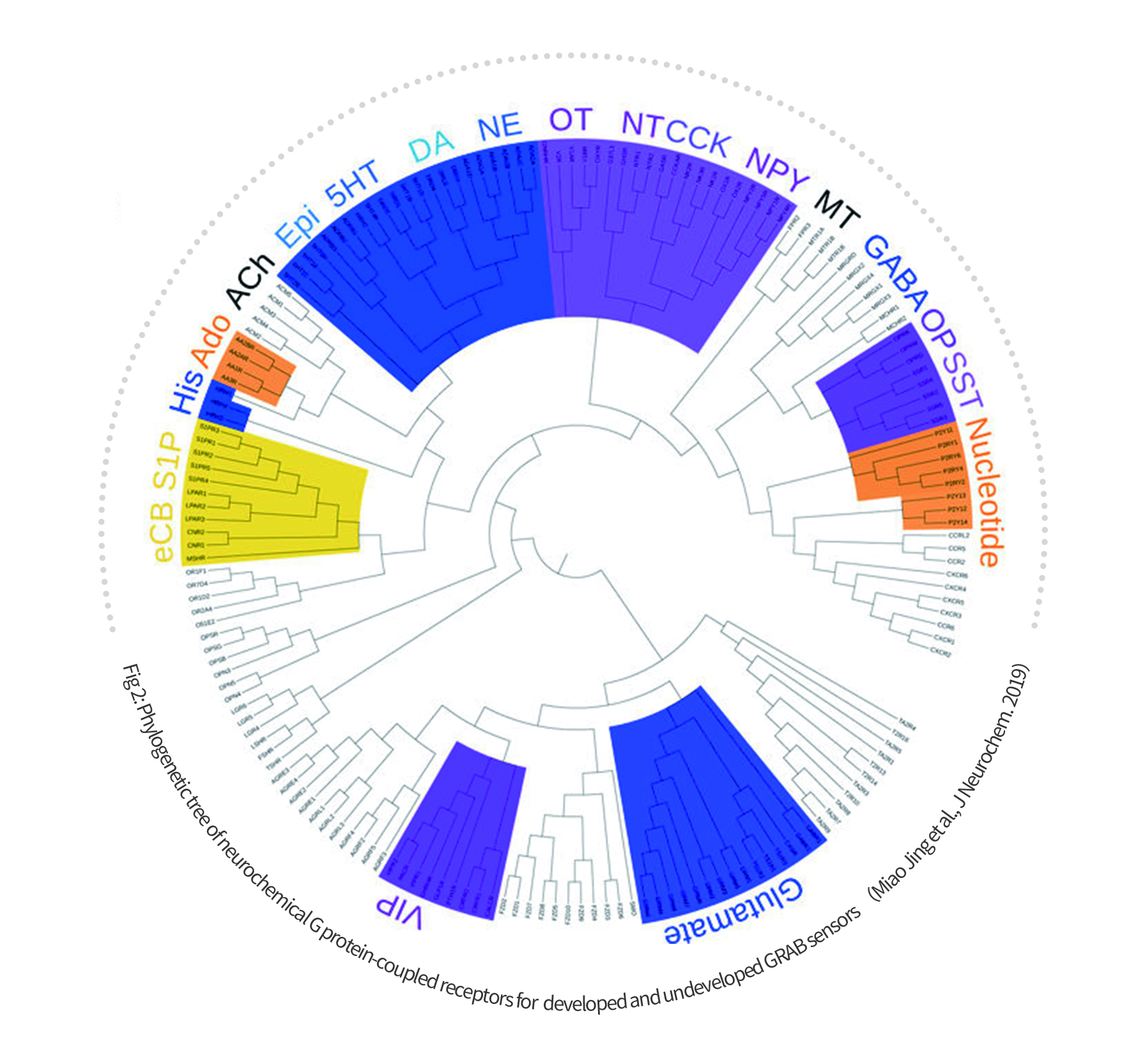
WZ Biosciences is the sole distributor of GRAB sensors in AAV viruses for in vivo monitoring of neurochemicals (Fig 2).
Sensors available include those for acetylcholine (ACh), dopamine (DA), NE, Serotonin (5-HT), adenosine (Ado), adenosine triphosphate (ATP), vasoactive intestinal peptide (VIP), cholecystokinin (CCK), endocannabinoid (eCB), neuropeptide Y (NPY), corticotropin-releasing factor (CRF), arginine vaso-pressin (AVP), histamine (His), somatostatin (SST), melatonin (MT), anandamide also known as N-arachidonoylethanolamine (AEA), oxytocin (OXT), Histamine (HA) and so on. Most of the GRAB sensors also have Cre-dependent and mutant versions. Some are also available in both cpGFP and cpRFP formats. More GRAB sensors are under development.
For all the published GRAB sensors, researchers can directly order from WZ Biosciences with discounted price (subsidized by the Li lab). For all the unpublished GRAB sensors, WZ Biosciences can help coordinate the purchase from the Li lab. Researchers can also directly communicate with the Li lab at yulonglilab2018@gmail.com.
4. FAQs
1) What is the suggested amount of viruses for injection?
Depending on the titer of each lot, we recommend 200-400nl per site of the non-diluted viruses.
2) Whether GRAB sensors will work in rats as they are in mice?
The GRAB sensors work well in rats. According to the published literature, these sensors can be used to detect neurotransmitter dynamics in many organisms, including Drosophila, zebrafish, mice and zebra finches.
3) Do you have the GRAB sensor AAVs with a ubiquitous promoter or other tissue-specific promoter?
Please refer to the list of tissue-specific promoters below to choose your interested promoter. Please contact us if you do not find the promoter of interest.
|
Tissue
|
Promoter Name
|
Size
|
Description
|
Aplication
|
|
CNS
|
hSyn
|
471bp
|
Human synapsin I promoter
|
Specific in neuron
|
|
CamKIIa
|
1.2kb
|
Mouse α-calcium-calmodulin dependent kinase II promoter
|
Specific in excitatory neurons in the neocortex and hippocampus
|
|
c-fos
|
1.7kb
|
mouse c-fos gene promoter
|
Specific in excitatory neurons
|
|
Mecp2
|
230bp
|
mouse methyl CpG binding protein 2 promoter
|
Specific in neuron, short
|
|
NSE
|
1.3kb
|
Mouse enolase promoter
|
Specific in neuron
|
|
Somatostat(SST)
|
1.2kb
|
Human somatostatin I gene
|
Specific in GABAergic Inhibitory neuron subtype (SST)
|
|
TH
|
2.5kb
|
Rat tyrosine hydroxylase gene promoter
|
Specific in dopaminergic neurons
|
|
GFAP
|
2.0kb
|
Human glial fibrillary acidic protein promoter
|
Specific in astrocyte
|
|
GFAP104
|
845bp
|
Human glial fibrillary acidic protein promoter
|
Specific in astrocyte
|
|
GfaABC1D(truncated GFAP)
|
681bp
|
Human glial fibrillary acidic protein promoter
|
Specific in astrocyte
|
|
ALDH1L1
|
1.3kb
|
Human aldehyde dehydrogenase 1 family member L1 promoter
|
Specific in astrocyte in thalamus
|
|
MBP
|
1.3kb
|
Human myelin basic protein promoter
|
Specific in oligodendrocyte
|
|
Liver
|
ALB
|
2.4kb
|
Mouse albumin promoter
|
Specific in liver
|
|
TBG
|
460bp
|
Human thyroxine binding globulin promoter
|
Specific in liver
|
|
ApoEHCR-hAAT
|
1.3kb
|
Chimeric promoter of the hepatocyte control region (HCR) from the APOE gene and the
human α1-antitrypsin promoter (hAAT)
|
Specific in liver
|
|
Heart
|
aMHC
|
0.4kb
|
Mouse myosin heavy chain alpha promoter
|
Specific in cardiomyocyte
|
|
cTNT+intron
|
0.7kb
|
Chicken cardiac troponin T promoter
|
Specific in cardiomyocyte
|
|
Eye
|
Rep65
|
0.7kb
|
Mouse retinal pigment epithelium 65 promoter
|
Specific in retinal pigment epithelium(RPE)
|
|
VMD2 promoter
|
0.65bp
|
Human vitelliform macular dystrophy-2 promoter
|
Specific in retinal pigment epithelium(RPE)
|
|
Pancreas
|
Insulin
|
0.85kb
|
mouse insulin promoter
|
Specific in pancreatic β cells
|
|
PDX1
|
2.7kb
|
Mouse pancreatic and duodenal homeobox 1 promoter
|
Specific in pancreatic β cells
|
|
Blood vessel
|
SM22a
|
0.45kb
|
Mouse SM22 alpha promoter
|
Specific in vascular smooth muscle cell
|
|
ICAM2
|
0.15kb
|
Human Intercellular Adhesion Molecule 2 promoter
|
Specific in endothelial cell
|
|
CD68
|
0.7kb
|
human CD68 promoter
|
Specific in monocytes and macrophages
|
|
F4/80
|
1.2kb
|
mouse F4/80 gene promoter
|
Specific in macrophages
|
|
Muscle
|
MCK
|
1.3kb
|
mouse muscle creatine kinase gene promoter
|
Specific in muscle
|
|
3×enhancer McK
|
728bp
|
modified mouse muscle creatine kinase gene promoter
|
Specific in muscle
|
|
Kidney
|
NPHS1
|
1.2kb
|
mouse Nephrin gene promoter
|
Specific in kidney
|
4) How much viruses are in one vial?
We provide the GRAB sensor AAVs in 50ul aliquots.
5) There are many AAV serotypes. Have you packaged any of GRAB sensors with other serotypes, such as a retrograde AAV serotype?
For most of GRAB sensors, we package them in the AAV9 serotype. If the retrograde serotype is required, we offer custom packaging services.
5. Research Background
One of the main focuses of neuroscience studies at cellular and molecular levels is to understand how neurons are connected and communicate with each other. Communication between neurons at the synapse depends on chemical transmissions. There are two types of chemical messenger at the synapse, neurotransmitters and neuromodulators. Neurotransmitters diffuse across the synaptic cleft to bind to ligand-gated ion channels on the postsynaptic cells, while neuromodulators bind to presynaptic and postsynaptic GPCRs. Upon the binding of neurotransmitters such as glutamate and GABA, ligand-gated ion channels rapidly depolarize or hyperpolarize the postsynaptic neurons and directly regulate the activity of these neurons. For most neuromodulators, theirs binding to GPCRs cascades serial events within the presynaptic and postsynaptic neurons and generate slow but long-term changes in these neurons.
To manipulate and monitor the communication between neurons, various methods have been established and many tools (e.g. delivery by AAVs) have been developed. Nowadays, optogenetics and chemical genetics tools are used to directly control the generation of presynaptic action potentials. Calcium indicators are used to monitor the signal propagation within the postsynaptic neurons. However, the techniques to monitor the releasing of neurotransmitters and neuromodulators with high sensitivity, specificity and precise spatial resolution are not well developed.
6. References
[1] Wan, J., et al. (2021). "A genetically encoded sensor for measuring serotonin dynamics." Nat Neurosci.
[2] Sun, F., et al. (2020). "Next-generation GRAB sensors for monitoring dopaminergic activity in vivo." Nat Methods 17(11): 1156-1166.
[3] Jing, M., et al. (2020). "An optimized acetylcholine sensor for monitoring in vivo cholinergic activity." Nat Methods 17(11): 1139-1146.
[4] Peng, W., et al. (2020). "Regulation of sleep homeostasis mediator adenosine by basal forebrain glutamatergic neurons." Science 369(6508).
[5] Jing, M., et al. (2019). "G-protein-coupled receptor-based sensors for imaging neurochemicals with high sensitivity and specificity." J Neurochem 151(3): 279-288.
[6] Feng, J., et al. (2019). "A Genetically Encoded Fluorescent Sensor for Rapid and Specific In Vivo Detection of NE." Neuron 102(4): 745-761 e748.
[7] Sun, F., et al. (2018). "A Genetically Encoded Fluorescent Sensor Enables Rapid and Specific Detection of Dopamine in Flies, Fish, and Mice." Cell 174(2): 481-496 e419.
[8] Jing, M., et al. (2018). "A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies." Nat Biotechnol 36(8): 726-737.